PUBLICIDAD

Estudios actuales han mostrado que el bloqueo de los receptores mGlu5 perjudica el aprendizaje espacial evaluado con una tarea en el laberinto acuático con ratones, sugiriendo una implicación de estos receptores en el aprendizaje y la memoria. En un intento por esclarecer la participación de los receptores mGlu5 en los procesos de aprendizaje, hemos examinado los efectos de 2-metil-6-(feniletinil)piridina (MPEP; 10 mg/kg, i. p. ), un antagonista selectivo de los receptores de glutamato mGlu5, en el aprendizaje espacial con ratones macho de la cepa OF. 1. Para ello, empleamos una tarea de aprendizaje en el laberinto de hoyos adaptada a ratón, que nos permite la valoración simultánea de la memoria de trabajo (MT) y memoria de referencia (MR) espacial. El laberinto de hoyos consiste en un campo abierto que consta de 16 hoyos.
Durante los ensayos, la misma configuración de 4 hoyos reforzados permanece constante a lo largo del entrenamiento. Los animales deberían aprender a visitar sólo las localizaciones reforzadas y hacerlo una sola vez, recordando la lista de lugares ya visitados para evitar las re-visitas. Nuestros resultados mostraron que los ratones adquirieron fácilmente esta tarea a los 5 días (4 ensayos por día). El último día, 30 minutos antes del entrenamiento, los animales fueron tratados con solución salina (grupo control) o con MPEP. No encontramos diferencias significativas entre ambos grupos. Estos hallazgos indican que la administración aguda de MPEP (10 mg/kg) no afecta la MT y MR espacial en ratones macho. Se hacen necesarios otros estudios con mayores dosis del fármaco y otras situaciones de prueba para confirmar estos resultados.
Efectos de la administración de MPEP, un antagonista selectivo de los receptores de glutamato mGLU5, sobre la memoria espacial en ratones.
(Effects of MPEP administration, a selective antagonist of mGLU5 receptors, on a spatial memory task in mice. )
E. Blanco(1); M. Martin-Lopez (1); M. Cavas (1); J. L. Arias (2); J. F. Navarro(1).
(1) Department of Psychobiology, Faculty of Psychology, University of Malaga, Spain
(2) Laboratory of Psychology, Faculty of Psychology, University of Oviedo, Spain
PALABRAS CLAVE: Receptor metabotrópico de glutamato, mGluR5, MPEP, Espacial, aprendizaje, memoria, Ratón
(KEYWORDS: Metabotropic glutamate receptor, mGluR5, MPEP, Spatial, Learning, Memory, Mice. )
Resumen
Estudios actuales han mostrado que el bloqueo de los receptores mGlu5 perjudica el aprendizaje espacial evaluado con una tarea en el laberinto acuático con ratones, sugiriendo una implicación de estos receptores en el aprendizaje y la memoria. En un intento por esclarecer la participación de los receptores mGlu5 en los procesos de aprendizaje, hemos examinado los efectos de 2-metil-6-(feniletinil)piridina (MPEP; 10 mg/kg, i. p. ), un antagonista selectivo de los receptores de glutamato mGlu5, en el aprendizaje espacial con ratones macho de la cepa OF. 1. Para ello, empleamos una tarea de aprendizaje en el laberinto de hoyos adaptada a ratón, que nos permite la valoración simultánea de la memoria de trabajo (MT) y memoria de referencia (MR) espacial. El laberinto de hoyos consiste en un campo abierto que consta de 16 hoyos. Durante los ensayos, la misma configuración de 4 hoyos reforzados permanece constante a lo largo del entrenamiento. Los animales deberían aprender a visitar sólo las localizaciones reforzadas y hacerlo una sola vez, recordando la lista de lugares ya visitados para evitar las re-visitas. Nuestros resultados mostraron que los ratones adquirieron fácilmente esta tarea a los 5 días (4 ensayos por día). El último día, 30 minutos antes del entrenamiento, los animales fueron tratados con solución salina (grupo control) o con MPEP. No encontramos diferencias significativas entre ambos grupos. Estos hallazgos indican que la administración aguda de MPEP (10 mg/kg) no afecta la MT y MR espacial en ratones macho. Se hacen necesarios otros estudios con mayores dosis del fármaco y otras situaciones de prueba para confirmar estos resultados.
Abstract
Recent studies have found that the blockade of the mGlu5 receptors impairs spatial learning tested in a Water maze task in mice, suggesting a role for these receptors in learning and memory. In an attempt to clarify the implication of mGLU5 receptors in spatial learning processes, we examined the effects of 2-methyl-6-(phenylethylnyl) pyridine (MPEP; 10 mg/kg, i. p. ), a selective antagonist of the mGlu5 receptors, on spatial learning in male mice of the OF. 1 strain. For this purpose, we have adapted to mice a holeboard-learning task, which allows the simultaneous assessment of spatial working (WM) and reference memory (RM) performance. The hole-board apparatus consists of an open-field with a 16-hole floor inserted. Across trials, a configuration of 4 baited holes with a food pellet was unchanged along training. Animals must to learn to visit only baited locations and to visit them only once, remembering a list of places already visited in order to avoid revisits. Our results showed that mice readily acquired this task within 5 days (4 trials per day). The last day, 30 minutes before training, animals were treated either with saline solution (control group) or with MPEP. No significant differences were found between both groups. These findings indicate that acute administration with MPEP (10 mg/kg) did not affect spatial WM and RM in male mice. Further studies with higher doses of the drug and other test situations are needed to confirm these results.
Introducción
El glutamato es el neurotransmisor excitador más abundante del sistema nervioso. Su acción tiene lugar a través de receptores de glutamato ionotrópicos (iGluRs), AMPA, NMDA y Kainato, y receptores de glutamato metabotrópicos (mGluRs). Los mGluRs forman una familia con ocho subtipos de receptores, divididos en tres grupos, grupo I: mGluR1 y mGluR5; grupo II: mGluR2 y mGluR3, y grupo III: mGluR4, mGluR6, mGluR7 y mGluR8. Los mGluRs del grupo I, actúan vía proteínas G acoplándose a fosfolipasa C (1). De hecho, la activación de este fosfolípido es necesaria para la inducción de PLP (Potenciación a Largo Plazo) en la región hipocampal, relacionada con procesos de aprendizaje asociativo entre estímulos espaciales y su contexto ambiental (2). Así, se ha observado que tanto por el bloqueo con antagonistas selectivos como por la supresión genética de estos receptores, la activación endógena de mGluR1 y mGluR5 es necesaria para el condicionamiento contextual y el aprendizaje de lugar (3-6).
Concretamente, el mGluR5 ha sido relacionado con procesos de plasticidad sináptica, aprendizaje y memoria. El MPEP (2-metil-6-(feniletinil)piridina) es un potente y selectivo antagonista no-competitivo del mGluR5 (7), cuyo desarrollo ha facilitado la comprensión del papel de este receptor en el sistema nervioso central. La localización del mGluR5 es fundamentalmente postsináptica, presentando una alta densidad en las regiones de hipocampo, corteza, cerebelo y sistema olfatorio (8). Su estimulación posibilita la liberación de calcio desde el almacén intracelular y la activación de proteína Kinasa C. Recientemente, se ha descrito que el mGluR5 puede funcionar como elemento clave a la hora de determinar una adecuada ejecución en un aprendizaje de orientación espacial entre diferentes cepas de rata. Estas diferencias de plasticidad sináptica vienen causadas por un bloqueo diferencial de la PLP tras la administración de MPEP, alterando los niveles de expresión del mGluR5 en la región CA1 hipocampal (9).
Numerosos estudios en modelos animales de emoción han descrito con detalle que los antagonistas del mGluR5 ejercen un claro efecto ansiolítico (para revisión, ver 10, 11, 12) y antiagresivo (13). Sin embargo, su acción sobre las funciones cognitivas no ha sido suficientemente explorada, aun siendo un área de interés, ya que los mGluR5s presentan altos niveles de expresión en la formación hipocampal (14).
Los efectos de la administración de MPEP en varios paradigmas conductuales muestran que los mGluR5s juegan un papel crítico en tareas de aprendizaje aversivas y aprendizaje espacial hipocampo-dependiente. No obstante, el MPEP no ejerce ningún efecto en otras formas de aprendizaje. Por ejemplo, parece existir un deterioro del aprendizaje en pruebas como el laberinto acuático de Morris, el radial y tareas de condicionamiento del miedo contextual, pero no en tareas de condicionamiento guiado (para revisión, ver 15). Por ello, los resultados de algunos trabajos existentes pueden resultar contradictorios. La eliminación del mGluR5 en ratones Knockout parece perjudicar la realización de pruebas cognitivas (5). De hecho, la administración de MPEP puede deteriorar la cognición, particularmente en la realización de algunas tareas de memoria de trabajo espacial (16, 17). No obstante, existen numerosos trabajos que muestran como el MPEP no ejerce efectos claros en la adquisición de tareas espaciales (18-21). En definitiva, los resultados existentes van a presentar diferencias notables dependiendo de múltiples factores, como la vía de administración de la droga, la dosis empleada y el tipo de prueba utilizada, sobretodo dependiendo de la carga emocional y/o puramente cognitiva de la tarea.
El objetivo de este trabajo fue estudiar la influencia de la administración aguda de MPEP sobre el aprendizaje espacial en una tarea de memoria de trabajo (MT) y memoria de referencia (MR) en el laberinto de hoyos en ratones macho.
Material y Métodos
Animales
Se emplearon 19 ratones macho adultos de la cepa OF. 1 (25–30 gr. ) obtenidos del bioterio central de la Universidad de Málaga. Los animales fueron alojados bajo condiciones normales de luz (luz blanca de 20:00 a 8:00), temperatura (21°C) y humedad (65 5%) constantes, con comida y agua ad libitum, excepto durante los días de habituación y entrenamiento, donde se les privó de comida la noche anterior a la realización de la tarea. El acceso libre a la comida se restauró una hora después de finalizar el último ensayo de cada día.
Los animales fueron asignados aleatoriamente a dos grupos: grupo Control Salino (n = 9) y grupo MPEP (10 mg/kg, i. p. ) (n = 10)
El uso y manipulación de los animales fue realizada respetando las normas éticas para la experimentación animal según la Directiva 86/609/CEE del Consejo (24 de Noviembre de 1986), cumpliendo las disposiciones legales, reglamentarias y administrativas de los Estados miembros de la Unión Europea sobre protección de los animales utilizados para experimentación y otros fines científicos y, legislados en nuestro país, mediante el Real Decreto 1201/2005 (10 de octubre de 2005).
Laberinto de Hoyos
El laberinto de hoyos consiste básicamente en un campo abierto cuadrado (40 X 40 cm. ) cercado por paredes laterales transparentes (25 cm. ), con 16 orificios (diámetro 3, 5 cm. ) donde se coloca un único reforzador de comida (≈ 20 mg. ) en 4 de ellos. Para controlar las pistas olfativas, todos los hoyos poseen comida en un doble fondo evitando el acceso directo a los mismos. Antes de que cada ratón fuera evaluado, se limpiaba la superficie y las paredes del laberinto con una mezcla de agua y alcohol al 5%.
El laberinto se colocó en una habitación aislada que disponía de diferentes pistas colgadas en las paredes que podrían ser empleadas para facilitar el proceso de orientación espacial en la búsqueda de los reforzadores. Todas las pruebas se realizaron siempre en horario de mañana (9:00- 14:00).
Tarea de aprendizaje espacial
La tarea de aprendizaje en el laberinto de hoyos nos permite valorar de manera simultánea memoria de trabajo (MT) y memoria de referencia (MR) espacial, evaluando la capacidad de los animales para recordar qué 4 localizaciones de las 16 existentes contienen comida.
El proceso comenzó tras manipular a los animales y habituarlos al reforzador en sus cajas para evitar reacciones neofóbicas durante 2 días. El procedimiento comportamental se inició con 2 días de habituación al laberinto en ensayos de 15 min. , donde se permitió la exploración libre de la arena y el acceso a los reforzadores distribuidos por todos los hoyos y la superficie de la caja. A continuación, empezó la fase de aprendizaje con 5 días consecutivos de entrenamiento y 4 ensayos diarios. 30 minutos antes de comenzar el último día de aprendizaje (día 6), los animales fueron inyectados con MPEP (Laboratorios Tocris) diluido en suero salino fisiológico a una dosis de 10 mg/kg. Tanto el fármaco como el suero salino fueron inyectados intraperitonealmente en un volumen de 10 ml/kg.
Cada ensayo comenzó colocando al animal de cara a cada una de las 4 esquinas de la arena, para iniciar la búsqueda de 4 reforzadores (localización espacial constante) entre los 16 hoyos, y terminó cuando el animal había encontrado todos los reforzadores o habían transcurrido 180 sg. de tiempo máximo. Durante el aprendizaje, se mantuvo la misma configuración de hoyos reforzados a lo largo de todo el entrenamiento. Los animales debían aprender a visitar únicamente los hoyos reforzados y hacerlo una sola vez. Así, recordarían la lista de lugares ya visitados evitando cometer errores al acceder a hoyos que ya no contienen ningún refuerzo (re-visita) porque han sido recolectados con anterioridad.
De esta forma, podemos operativizar las medidas de memoria de referencia (MR) y memoria de trabajo (MT) espacial, mediante el cálculo de unos índices. MR, expresa el número de visitas a la configuración de hoyos reforzados (visitas y re-visitas) como proporción del número total de visitas a todos los hoyos (reforzados y no-reforzados). MT, esta medida representa la ratio de todas las visitas a la configuración de hoyos reforzados (visitas y re-visitas) que han sido reforzadas con comida. Para ello grabamos todo el proceso con una videocámara y analizamos las imágenes a posteriori contabilizando la frecuencia de visita a los hoyos en todas sus posibilidades y las latencias de búsqueda hasta alcanzar los reforzadores.
Resultados
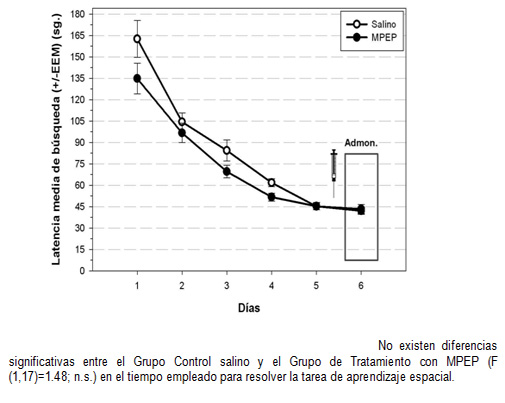
Realizamos un ANOVA de medidas repetidas que muestra un efecto significativo en los días (F (4, 68)=43. 75; p≤ 0. 001), pero no entre los grupos (F (1, 17)=1. 53; n. s. ), ni en la interacción días x grupos (F (4, 68)=0. 69; n. s. ). Estos resultados muestran que los ratones de ambos grupos adquirieron progresivamente la tarea durante los 5 primeros días de entrenamiento (Gráfica 1). El último día (día 6), 30 minutos antes del entrenamiento, los animales fueron inyectados con solución salina (grupo control) o con MPEP. Tampoco encontramos diferencias significativas entre ambos grupos (F (1, 17)=1. 48; n. s. ) en la latencia de búsqueda de los reforzadores, al incluir este último día de test. Ésto nos indica que la administración aguda de MPEP (10 mg/kg) no afectó el aprendizaje de la tarea en ratones macho.
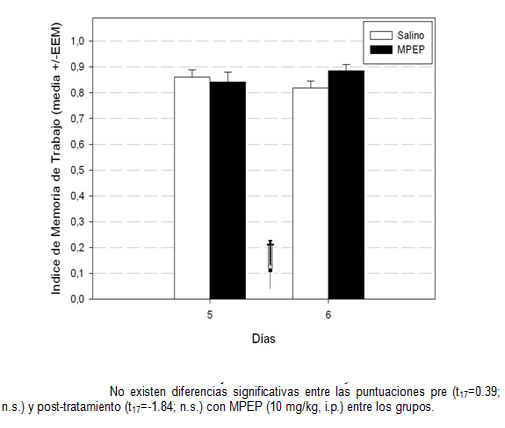
En cuanto a los índices de memoria, podemos observar que no existen diferencias significativas en MR entre los grupos ni antes (t17=-1. 27; n. s. ), ni después (t17=-0. 95; n. s. ) del tratamiento con MPEP. Lo mismo ocurre con la MT, no se muestran diferencias entre grupo control y grupo con MPEP pre (t17=0. 39; n. s. ) y post-tratamiento (t17=-1. 84; n. s. ). Es decir, que la administración aguda de MPEP a una dosis de 10 mg/kg, no afecta la MT y MR en el aprendizaje espacial realizado en el laberinto de hoyos.
Gráfica 1: Latencia media de búsqueda durante el entrenamiento
Gráfica 2: Índice de memoria de Referencia durante los días 5 y 6 del entrenamiento en el laberinto de 16 hoyos
Gráfica 3: Índice de memoria de Trabajo durante los días 5 y 6 del entrenamiento en el laberinto de hoyos
Discusión
Los resultados del presente trabajo muestran que el antagonista del mGluR5, MPEP, a una dosis aguda de 10 mg/kg, no perjudicó el aprendizaje espacial de ratones macho en el laberinto de hoyos. La inyección de una dosis de MPEP 30 minutos antes de la realización de la tarea el último día de test no modificó la ejecución de los roedores en MR y MT espacial.
Resultados similares han sido obtenidos empleando inyecciones con dosis entre 1-10 mg/kg (i. p. ) antes de cada sesión de entrenamiento en una tarea espacial de triple elección reforzada (18). Independientemente de la dosis utilizada, la administración de MPEP (i. p. ) previa al entrenamiento, no causa efectos en la adquisición de esta tarea de aprendizaje espacial de búsqueda de reforzadores. También se han probado dosis mayores, con rangos entre 3-30 mg/kg administradas por vía oral (p. o. ) antes del ensayo de prueba, pero con idénticos resultados donde los grupos tratados ejecutan la tarea de forma similar a los animales controles en el laberinto acuático de Morris (22). En este mismo test, tratando a animales adultos con 1 mg/kg por vía intravenosa (i. v. ) 20 minutos antes de la prueba, tampoco se han observado efectos que permitan afirmar que el MPEP ejerce un papel importante en el aprendizaje espacial (19). Como excepción a estos resultados, encontramos que la inyección subcutánea (s. c. ) de MPEP en un rango de dosis entre 0, 63 y 10 mg/kg, produce un leve d¨¦ficit en la adquisición del aprendizaje en la piscina, pero sólo con la dosis más alta del tratamiento (20).
En otras pruebas de memoria como la tarea de reconocimiento de objetos, la inyección de MPEP (6 mg/kg, i. p. ) 30 minutos antes o inmediatamente después del entrenamiento no interfiere en el recuerdo del objeto familiar (23). Tampoco se produce ningún efecto cuando se administra MPEP (3-10 mg/kg, i. c. ) en la corteza perirrinal, con una demora de reconocimiento de 15 minutos (memoria a corto plazo) o 24 horas (memoria a largo plazo) después (24).
A su vez, se ha observado que dentro del grupo I, los mGluR1 pueden tener una mayor implicación que los mGluR5 en ciertos procesos cognitivos. Por ejemplo, parece que existen efectos diferenciales entre el antagonista del mGluR1, 4-CPG, y el antagonista del mGluR5, MPEP. Se ha visto que mientras que 4-CPG bloquea la adquisición a largo plazo en la tarea espacial de triple elección reforzada, el MPEP no tiene efectos sobre la tarea de aprendizaje (18). Otro antagonista del mGluR1, JNJ16259685, también produce un potente efecto de bloqueo sobre la adquisición y retención del aprendizaje espacial, que comparado con la administración de MPEP (s. c. ) se reduce a un efecto mínimo y sólo con la dosis más alta de las ensayadas (0, 63-10 mg/kg) (20).
Hasta el momento, los resultados revisados junto a los datos presentados en nuestro trabajo, nos permiten observar que el efecto de la administración sistémica de MPEP no es suficiente para afirmar que el mGluR5 desarrolla un papel importante en la modulación de los procesos de memoria.
En cambio, existen otros trabajos que han empleado otras vías de administración de este fármaco. Así, sabemos que la inyección intracerebroventicular (i. c. v) de MPEP 30 minutos antes del entrenamiento en una tarea de alternancia espacial en el laberinto en Y, no provocó efectos en la adquisición de la prueba, aunque si ejerció un severo déficit de retencón al día siguiente (4, 17). La administración inmediata post-entrenamiento no afectó la respuesta de alternancia (17), pero la infusión pre-entrenamiento perjudicó el aprendizaje en la prueba de alternancia espacial (4, 17). También conocemos como las administraciones repetidas con esta forma de administración (i. c. v) durante 10 días, causaron un déficit importante en memoria de referencia y memoria de trabajo espacial en el laberinto radial (16, 25). Algunos de estos trabajos que estudian los efectos de los antagonistas del mGluR5 en la cognición han sido realizados mediante administración intracerebral (i. c. ), pudiendo provocar efectos contrapuestos a la administración sistémica de fármacos (i. p. , o. p. , i. v. ). De lo visto hasta ahora, parece que mientras que la administración i. c. /i. c. v. de MPEP podría perjudicar la realización de tareas de memoria espacial, la administración sistémica no provocaría efectos negativos sobre la adquisición y el recuerdo de una tarea espacial. Las razones para estas discrepancias no están todavía claras, pero junto a la influencia de las vías de administración podemos apuntar a que diferencias en las dosis utilizadas, la concentración del fármaco en el sistema nervioso y el tipo de paradigma conductual empleado para la evaluación, podrían ser aspectos muy relevantes a la hora de extraer conclusiones.
No podemos obviar algunos hechos destacables sobre los efectos del MPEP en los procesos de plasticidad sin¨¢ptica, como la PLP hipocampal. Balschun y Wetzel (2002), mostraron que la inyección i. c. v. de MPEP (13, 8 ¦Ìgr. ) previa a la tetanización, producía un deterioro de la PLP (7 minutos), con una disminución de la potenciación que se prolongaba durante las 6 horas restantes (17). De hecho, la administración i. c. v. repetida de MPEP con 30 minutos de antelación a la tetanización, perjudicó la PLP en el giro dentado causando un deterioro del aprendizaje espacial (16, 25). Nuestros datos no coinciden con los resultados de estos trabajos; sin embargo, considerando las vías de administración podrían producirse diferentes efectos debidos a la exposición al fármaco, que variarían entre la concentracón directamente inyectada (i. c. v. ) y la concentración total en cerebro (i. p. ). Por ejemplo, una dosis alta de MPEP (13, 8 ¦Ìgr. ) directamente inyectada en ventrículo podría movilizar no sólo receptores mGlu5 sino también receptores NMDA (26, 27), que también afectarían la PLP. Algunos autores podrían achacar la ausencia de efectos conductuales con la administración sistémica de MPEP a una concentración insuficiente del fármaco disuelta en cerebro, pero se ha visto que inyecciones similares afectan la actividad de las neuronas CA1 del hipocampo (7) e influencian la adquisición en el aprendizaje de tareas punitivas (28). De hecho, estudios in vivo han demostrado que dosis de MPEP de 10 mg/kg (i. p. ) logran una ocupación total de los receptores mGluR5, que se mantiene al máximo durante 1 hora reduciéndose a lo largo de un período de 4 horas (29). Por tanto, se debe considerar que el MPEP es un fármaco activo a nivel cerebral después de inyecciones sistémicas (7) y, en consecuencia, podría ser inyectado periféricamente para evaluar sus efectos conductuales tal y como hemos visto.
De otro modo, conocemos que los antagonistas del mGluR5 poseen un marcado perfil ansiolítico (10-12). Así, sabemos que la administración oral de MPEP induce un sólido efecto en diferentes tareas emocionales (test de Geller-Seifter, condicionamiento de la respuesta emocional, test de Vogel, condicionamiento operante de palanca demorado) comparables a los efectos producidos por otros ansiolíticos, como el diazepam; pero a diferencia de éste, no provoca una afectación cognitiva (memoria espacial) (22). Se ha comprobado que a dosis ansiolíticas efectivas, tanto en administración aguda como sub-aguda, no se produce un deterioro cognitivo de la conducta cuando se evalúa a los animales en pruebas dependientes de un reforzador positivo (condicionamiento operante de palanca, laberinto de hoyos -este trabajo-) o negativo (laberinto acu¨¢tico de Morris) (22).
Además, parece que existen algunas diferencias en la utilización de MPEP frente al uso de otros antagonistas como el bloqueador del mGluR1, 4-CPG. Se ha observado que el 4-CPG perjudica la adaptación espontánea en la exploración de un campo abierto, al contrario que el MPEP con el que se desarrolla un patrón de exploración similar a los animales del grupo control (18). Otro antagonista del mGluR1, JNJ16259685, perjudica el aprendizaje a dosis por debajo de las requeridas para ejercer propiedades ansiolíticas, mientras que el MPEP muestra efectos ansiolíticos a dosis a las que no se producen déficits de aprendizaje (20, 30). En este caso, la diferencia entre ambos antagonistas glutamatérgicos viene dada por una mayor ocupación del mGluR a nivel central por parte del MPEP (20). Esto podría significar una ventana de seguridad terapéutica favorable para el antagonista del mGluR5, aunque no podemos olvidar que el MPEP también interactúa con transportadores de la noradrenalina con baja afinidad, que podrían contribuir a potenciar sus propiedades ansiolíticas (31). El hecho de que el MPEP ejerza un perfil ansiolítico estable similar al de otros ansiolíticos clásicos y, al contrario que éstos, no perjudique las capacidades cognitivas a dosis ansiolíticas (diazepam) hace pensar que podría ser usado en un futuro con fines clínicos (22). Esto último, unido a una nula afectación de la actividad motora espontánea, sin limitación de dosis para la sedación y una escasa interacción con el alcohol, podría sugerir que los antagonistas del mGluR5 puedan proporcionar un nuevo y eficaz tratamiento para los trastornos de ansiedad con un perfil de efectos mejorado sobre algunas benzodiacepinas.
El desarrollo actual de moduladores alostóricos positivos y negativos de los mGluR5s debería proveer de nuevas herramientas farmacológicas para incrementar nuestro conocimiento sobre los procesos fisiológicos y patofisioógicos de estos receptores, facilitando nuevas investigaciones para probar los mGluR5s como dianas terapeéticas en el tratamiento de desórdenes psicológicos y neurológicos.
Referencias
1. Conn PJ, Pin JP. Pharmacology and functions of metabotropic glutamate receptors. Annu Rev Pharmacol Toxicol 1997; 37:205-37.
2. Morris RG, Frey U. Hippocampal synaptic plasticity: role in spatial learning or the automatic recording of attended experience? Philos Trans R Soc Lond B Biol Sci 1997; 352(1360):1489-503.
3. Aiba A, Chen C, Herrup K, Rosenmund C, Stevens CF, Tonegawa S. Reduced hippocampal long-term potentiation and context-specific deficit in associative learning in mGluR1 mutant mice. Cell 1994; 79(2):365-75.
4. Balschun D, Wetzel W. Inhibition of group I metabotropic glutamate receptors blocks spatial learning in rats. Neurosci Lett 1998; 249(1):41-4.
5. Lu YM, Jia Z, Janus C, Henderson JT, Gerlai R, Wojtowicz JM, et al. Mice lacking metabotropic glutamate receptor 5 show impaired learning and reduced CA1 long-term potentiation (LTP) but normal CA3 LTP. J Neurosci 1997; 17(13):5196-205.
6. Rodrigues SM, Bauer EP, Farb CR, Schafe GE, LeDoux JE. The group I metabotropic glutamate receptor mGluR5 is required for fear memory formation and long-term potentiation in the lateral amygdala. J Neurosci 2002; 22(12):5219-29.
7. Gasparini F, Lingenhohl K, Stoehr N, Flor PJ, Heinrich M, Vranesic I, et al. 2-Methyl-6-(phenylethynyl)-pyridine (MPEP), a potent, selective and systemically active mGlu5 receptor antagonist. Neuropharmacology 1999; 38(10):1493-503.
8. Yu M. Recent developments of the PET imaging agents for metabotropic glutamate receptor subtype 5. Curr Top Med Chem 2007; 7(18):1800-5.
9. Manahan-Vaughan D, Braunewell KH. The metabotropic glutamate receptor, mGluR5, is a key determinant of good and bad spatial learning performance and hippocampal synaptic plasticity. Cereb Cortex 2005; 15(11):1703-13.
10. Spooren WP, Gasparini F, Salt TE, Kuhn R. Novel allosteric antagonists shed light on mglu(5) receptors and CNS disorders. Trends Pharmacol Sci 2001; 22(7):331-7.
11. Spooren W, Ballard T, Gasparini F, Amalric M, Mutel V, Schreiber R. Insight into the function of Group I and Group II metabotropic glutamate (mGlu) receptors: behavioural characterization and implications for the treatment of CNS disorders. In: Behav Pharmacol; 2003. p. 257-77.
12. Spooren W, Gasparini F. mGlu5 receptor antagonists: a novel class of anxiolytics? Drug News Perspect 2004; 17(4):251-7.
13. Navarro JF, Postigo D, Martin M, Buron E. Antiaggressive effects of MPEP, a selective antagonist of mGlu5 receptors, in agonistic interactions between male mice. Eur J Pharmacol 2006; 551(1-3):67-70.
14. Shigemoto R, Mizuno N. Metabotropic glutamate receptors-immunocytochemical and in situ hybridization analysis. In: O. P. Ottersen JS-M, editor. Handbook of chemical neuroanatomy. Amsterdam: Elsevier; 2000. p. 63-98.
15. Simonyi A, Schachtman TR, Christoffersen GR. The role of metabotropic glutamate receptor 5 in learning and memory processes. Drug News Perspect 2005; 18(6):353-61.
16. Manahan-Vaughan D, Schuetz K. Differential participation of metabotropic glutamate receptor mGlu1 and mGlu5 in spatial learning and hippocampal long-term potentiation in vivo. Neuropharmacology 2002; 43(Suppl):86.
17. Balschun D, Wetzel W. Inhibition of mGluR5 blocks hippocampal LTP in vivo and spatial learning in rats. Pharmacol Biochem Behav 2002; 73(2):375-80.
18. Petersen S, Bomme C, Baastrup C, Kemp A, Christoffersen GR. Differential effects of mGluR1 and mGlur5 antagonism on spatial learning in rats. Pharmacol Biochem Behav 2002; 73(2):381-9.
19. Car H, Stefaniuk R, Wisniewska RJ. Effect of MPEP in Morris water maze in adult and old rats. Pharmacol Rep 2007; 59(1):88-93.
20. Steckler T, Oliveira AF, Van Dyck C, Van Craenendonck H, Mateus AM, Langlois X, et al. Metabotropic glutamate receptor 1 blockade impairs acquisition and retention in a spatial Water maze task. Behav Brain Res 2005; 164(1):52-60.
21. Locchi F, Dall'Olio R, Gandolfi O, Rimondini R. Water T-maze, an improved method to assess spatial working memory in rats: Pharmacological validation. Neurosci Lett 2007; 422(3):213-6.
22. Ballard TM, Woolley ML, Prinssen E, Huwyler J, Porter R, Spooren W. The effect of the mGlu5 receptor antagonist MPEP in rodent tests of anxiety and cognition: a comparison. psychopharmacology (Berl) 2005; 179(1):218-29.
23. Christofferson GRJ, Simonyi A, Schachtman TR, Clement D, Skyjoth K, Bjerre V. Effects of mGluR1 and mGluR5 antagonism on exploratory locomotion and learning in a cross-maze and in an object recognition task. Physiology & Behavior 2008:Manuscript under review.
24. Barker GR, Bashir ZI, Brown MW, Warburton EC. A temporally distinct role for group I and group II metabotropic glutamate receptors in object recognition memory. Learn Mem 2006; 13(2):178-86.
25. Naie K, Manahan-Vaughan D. Regulation by metabotropic glutamate receptor 5 of LTP in the dentate gyrus of freely moving rats: relevance for learning and memory formation. Cereb Cortex 2004; 14(2):189-98.
26. O'Leary DM, Movsesyan V, Vicini S, Faden AI. Selective mGluR5 antagonists MPEP and SIB-1893 decrease NMDA or glutamate-mediated neuronal toxicity through actions that reflect NMDA receptor antagonism. Br J Pharmacol 2000; 131(7):1429-37.
27. Movsesyan VA, O'Leary DM, Fan L, Bao W, Mullins PG, Knoblach SM, et al. mGluR5 antagonists 2-methyl-6-(phenylethynyl)-pyridine and (E)-2-methyl-6-(2-phenylethenyl)-pyridine reduce traumatic neuronal injury in vitro and in vivo by antagonizing N-methyl-D-aspartate receptors. J Pharmacol Exp Ther 2001; 296(1):41-7.
28. Schachtman TR, Bills C, Ghinescu R, Murch K, Serfozo P, Simonyi A. MPEP, a selective metabotropic glutamate receptor 5 antagonist, attenuates conditioned taste aversion in rats. Behav Brain Res 2003; 141(2):177-82.
29. Anderson JJ, Bradbury MJ, Giracello DR, Chapman DF, Holtz G, Roppe J, et al. In vivo receptor occupancy of mGlu5 receptor antagonists using the novel radioligand [3H]3-methoxy-5-(pyridin-2-ylethynyl)pyridine). Eur J Pharmacol 2003; 473(1):35-40.
30. Steckler T, Lavreysen H, Oliveira AM, Aerts N, Van Craenendonck H, Prickaerts J, et al. Effects of mGlu1 receptor blockade on anxiety-related behaviour in the rat lick suppression test. psychopharmacology (Berl) 2005; 179(1):198-206.
31. Heidbreder CA, Bianchi M, Lacroix LP, Faedo S, Perdona E, Remelli R, et al. Evidence that the metabotropic glutamate receptor 5 antagonist MPEP may act as an inhibitor of the norepinephrine transporter in vitro and in vivo. Synapse 2003; 50(4):269-76.
IMPORTANTE: Algunos textos de esta ficha pueden haber sido generados partir de PDf original, puede sufrir variaciones de maquetación/interlineado, y omitir imágenes/tablas.


El uso de juegos de rol como herramienta terapéutica en la salud mental
Xavier Martin Alarcon
Fecha Publicación: 20/05/2024
")
Trastornos Cognitivos en la Salud Mental
Rita Carpanchai Colquillo
Fecha Publicación: 06/05/2024
")
EL USO DE JUEGOS DE ROL COMO HERRAMIENTA TERAPÉUTICA EN LA SALUD MENTAL
xavier Martin Alarcon
Fecha Publicación: 08/10/2023
Amnesia disociativa versus demencia vascular
Covadonga Huergo Lora et. al
Fecha Publicación: 18/05/2023
Impacto de la contención emocional en la comunidad educativa Gabriela Mistral en tiempos de Pandemia
Olga Viviana Chacón Hernández et. al
Fecha Publicación: 20/05/2022
")
“Mi saludo para ti Caballero, Caballero de París”: Un análisis sistémico de los mitos familiares.
ALBA ALBA PONS
Fecha Publicación: 09/05/2022